La mecanobiología estudia la interacción entre las señales mecánicas y los procesos biológicos que se producen en las células y tejidos.
La carga mecánica puede influir en la proliferación, diferenciación y metabolismo celular, por lo que tiene un papel crucial en el crecimiento, adaptación, regeneración y bioingeniería de los tejidos vivos.
La mecanobiología combina técnicas biológicas experimentales (modelos in vitro e in vivo) y técnicas computarizadas (modelos matemáticos e informáticos) para crear la interacción entre la mecánica y la biología.
Desde el punto de vista biomecánico, ningún tejido vivo puede compararse a otro material de ingeniería, ya que dichos tejidos presentan un continuo proceso de regeneración y remodelación que permite mejorar su estructura en función de los esfuerzos a los que es sometido (1).
Van der Meulen describió la mecanobiología esquelética como "la ciencia que estudia las fuerzas mecánicas que modulan la morfología y la adaptación estructural de los tejidos esqueléticos; es decir, hueso, cartílago, ligamento y tendón" (2).
Existen tres aspectos que han hecho avanzar enormemente la mecanobiología ósea en los últimos años:
- Los modelos informáticos de estructuras que permiten el análisis del efecto de fuerzas físicas en las complejas geometrías del hueso.
- La biología molecular que permite detectar la expresión de genes y la síntesis de proteínas tras la aplicación de diferentes fuerzas mecánicas.
- El avance de la tecnología de imagen, que permite identificar las características micro y nanoestructurales del tejido.
El método informático de elementos finitos (FE, por Finite Element) se trata de un eficaz análisis numérico para estudiar cuantitativamente el comportamiento de una estructura.
Este método ha sustituido a los iniciales procedimientos de fotoelasticidad, permitiendo no solo estudiar el predominio de la dirección del estrés sino también cuantificar la intensidad mecánica (3).
El objetivo de los FE es asimilar un supuesto comportamiento mecánico numérico al comportamiento biológico de las células de una estructura y de esta manera comparar los resultados de ambas evidencias de manera bidireccional.
A pesar del incremento en el conocimiento de los procesos celulares y moleculares que acontecen en la interfase hueso-implante, poco se conoce sobre el proceso por el cual las fuerzas mecánicas modulan el crecimiento óseo en esa interfase.
La mecanotransducción se encarga del estudio de la transmisión y distribución de las señales mecánicas, y su conversión en señales biológicas y químicas dentro de la célula (4).
La respuesta del hueso ante los estímulos mecánicos locales ya fue descrito por Wolff en 1892 (5) estableciendo la hipótesis de que "cada cambio de la forma y la función del hueso, o solo de la función, conlleva cambios definitivos en la arquitectura interna y la conformación externa, siguiendo leyes matemáticas".
Posteriormente se ha cuestionado la relación con posibles leyes matemáticas, pero lo que es incuestionable es que las cargas mecánicas no solo afectan el volumen del hueso sino también su organización interna.
Desde los trabajos iniciales de Frost (6,7) se ha aceptado de manera general que el hueso se renueva por si mismo a lo largo de la vida mediante las unidades multicelulares básicas (BMUs, por Bone Multicellular Units), que se forman de grupos de osteoblastos y osteoclastos que primero reabsorben el hueso y posteriormente lo van rellenando por un nuevo tejido.
El nuevo hueso formado se organiza en las osteonas del hueso compacto o en las hemi-osteonas o trabéculas del hueso trabecular alineándose según la dirección de las cargas dominantes (8) lo que sugiere que los gradientes de tensión local son un factor regulador de la forma en que se desarrolla el recambio óseo (9).
Basándose en la observación macroscópica, Wolff (5) estableció el primer planteamiento numérico de la estructura del hueso, con la descripción de la disposición perpendicular de las trabéculas en la cabeza del fémur.
Sin embargo, tanto el modelo de Wolf como el resto de los modelos numéricos posteriores no pueden ser tomados de manera categórica debido al número de factores no mecánicos que también influyen sobre el hueso (genéticos, nutricionales, hormonales, paracrinos, etc).
De esta manera se han establecido otros modelos en donde la disposición no es únicamente perpendicular, sino también con otras disposiciones (3).
Los modelos finitos tradicionales se han basado en niveles evidentes (macroscópicos) y en la premisa simplista del llamado "supuesto continuo" por el cual se presume un comportamiento similar en cada trabécula ósea.
El avance tecnológico de la microtomografia computarizada (micro-CT) y la creación de nuevos algoritmos ha permitido observar el comportamiento individual de una trabécula.
Parece que el estudio de elementos microestructurales y la formación posterior de elementos finitos entre 100 millones y 1 billón de elementos finitos establecerán unos modelos mas cercanos a la estructura real (3).
En este artículo se revisan los conceptos generales que se estudian en la mecanobiología y biomecánica de los hueso maxilares.
Se establecen las principales propiedades biomecánicas del hueso en las diferentes escalas de medición y determinados factores que influyen en la reacción del hueso perimplantario ante las cargas biomecánicas.
Propiedades biomecánicas del hueso (nivel macro, micro y nanométrico)
Hay que tener en cuenta que durante varios años se han establecido las propiedades biomecánicas del hueso basándose únicamente en la escala macrométrica (escala de centímetros o milímetros), valorando las diferencias estructurales visibles a simple vista que existen entre el hueso trabecular y el hueso cortical.
Sin embargo, hoy en día se puede estudiar la estructura del hueso a nivel orgánico, tisular, celular o molecular (10).
Las aproximaciones micro y nanoestructurales al hueso están reflejando las diferentes propiedades biomecánicas de los distintos tipos de tejido óseo que se pueden incluir tanto en el hueso trabecular como en el cortical, es decir el hueso reticular, el hueso de fibras paralelas y el hueso laminar.
De manera general, en el hueso, la resistencia (resistencia a la deformación) y la rigidez lo aporta la fase inorgánica, mientras que la tenacidad (resistencia a la fractura) lo aporta la fase orgánica (11).
Nivel macro y micrométrico (órgano/tejido/célula)
Teniendo en cuenta el carácter general anisotrópico no homogéneo del hueso, el hueso cortical se suele evaluar estructuralmente de manera imprecisa como un material isotrópico (con la misma propiedad biomecánica al ser medida en cualquier dirección), mientras que el hueso trabecular presenta mayor grado de anisotropía en función de las cargas que haya recibido en cada zona (1).
Si las propiedades son diferentes en las tres direcciones del espacio se habla de material ortotrópico, mientras que sin las propiedades son iguales en dos direcciones, se habla de isotropía transversal (12).
Las propiedades biomecánicas son variables entre el hueso cortical y trabecular principalmente porque el espacio poroso en el primero es del 5-10% mientras que en el segundo se encuentra entre 30-90% (10).
La morfología general del hueso, le permite tener una estructura rígida y ligera a la vez, ya que el hueso cortical facilita la rigidez mientras que las trabéculas internas minimizan el peso (1).
Las propiedades mecánicas se han valorado con diferentes métodos como son los ensayos mecánicos, la relación de microdureza Vickers con el módulo de elasticidad o las técnicas basadas en ultrasonidos.
Actualmente la herramienta más utilizada es la de creación de elementos numéricos o finitos mediante ordenador, que incluso puede añadir elementos reales a partir de imágenes de alta resolución.
De manera general, las propiedades biomecánicas del hueso van a depender de su contenido acuoso, de la porosidad (densidad) y del contenido mineral (1).
Hueso Cortical
Los valores obtenidos para el hueso cortical son menos dispersos que los encontrados en el hueso trabecular.
En general, el módulo de elasticidad se encuentra entre 15-20 GPa (13) la resistencia a la tensión se encuentra entre los 80-150 Mpa, y la resistencia a la compresión entre los 90-280 Mpa, debido principalmente al carácter anisotrópico del hueso (1).
Se ha observado que la cortical mandibular es anisotrópica, de tal forma que al igual que la diáfisis femoral, es mas rígida en la dirección longitudinal que en la radial o tangencial, teniendo en estas direcciones un 40-80% del valor de rigidez que tiene en la dirección longitudinal.
La resistencia a la compresión es de 200, 110 y 100 Mpa en la dirección longitudinal, tangencial y radial respectivamente (14).
Por otro lado hay que considerar que los valores de resistencia del hueso cortical son variables en función de la edad.
De esta manera la resistencia a la tensión máxima desciende desde los 120 Mpa con 30 años a 70 Mpa a los 100, mientras que el límite máximo de deformación cae del 3,3% a los 30 años al 1% a los 100 años.
De la misma manera se observan similares efectos en el hueso trabecular (15).
Se ha observado en hueso cortical femoral que a partir de los 35 años hay una disminución por década del módulo elástico de 0,35 GPa, de resistencia a la flexión de 6,25 Mpa, y de energía de fractura (tenacidad) de 0,03 Kj/m2.
El tamaño de las microgrietas también se ha observado aumentado con la edad con valores de 500 µm a los 35 años y de hasta 1 mm de longitud a los 92 años.
El aumento del número y tamaño de estas grietas se ha correlacionado positivamente con los resultados de tenacidad del hueso cortical (16).
Hueso Trabecular
Existen opiniones dispares en cuanto a establecer las propiedades biomecánicas del hueso trabecular.
Hay autores que lo consideran con un modulo de elasticidad similar al del hueso cortical (15-20 GPa) (13).
Otros autores, sin embargo le adjudican un valor de rigidez menor (0,75-10 GPa) (17).
En cualquier caso se han determinado diferentes valores de E para el hueso trabecular en función de la prueba de esfuerzo utilizada y las condiciones del hueso esponjoso (seco, fresco o congelado), aunque siempre con valores menores que el hueso cortical.
Los valores evidenciados son muy dispares incluso cuando los datos los emite un mismo autor aplicando el mismo ensayo a diferentes muestras (1,12).
McNamara y cols (18) desarrollaron un modelo sólido obteniendo 4 trabéculas de hueso vivo que posteriormente fueron procesadas con micro-CT y analizadas mediante elementos finitos.
Este modelo permite observar la reacción a cargas sobre un modelo con morfología real (incluyendo lagunas de reabsorción activas).
El estrés se observaba elevado en el fondo de las lagunas de reabsorción y aumentaría la actividad osteoclástica más allá de lo que inicialmente estaba previsto para renovar el hueso antiguo o dañado, y también evitan la distribución homogénea de la carga a lo largo de la trabécula.
Smit y Burger (19) establecieron la hipótesis que si la deformación baja (desuso) activa los osteoclastos, mientras que si la deformación aumenta (sobrecarga media) se activan los osteoblastos para rellenar la BMU.
Estos datos se observaron con unas medidas de estrés no mayores de 16 Mpa y de deformación no mayores de 3.074 µε y con modelos informáticos de trabécula ideales (sin obtenerlas de trabéculas in vivo).
Los datos de McNamara y cols (18) establecerían que aunque esto ocurriría habitualmente por debajo de 4.000 µε, existirían otros estímulos diferentes para activar a osteoclastos u osteoblastos cuando el estrés y las deformaciones son mayores de 4.000 µε.
Nivel nanométrico (molécula)
Durante muchos años se ha considerado que el hueso cortical y trabecular se consideraba un único material desde el punto de vista estructural molecular, y sus diferencias biomecánicas aparecerían en otras escalas debido a la diferencia de densidad.
Está hipótesis ha sido rechazado por la evidencia en diferentes estudios de variaciones biomecánicas intrínsecas en los dos tipos de hueso.
Si bien los componentes moleculares son idénticos, parece que la disposición de los mismos varía entre ambos influyendo en sus propiedades biomecánicas.
Se han observado valores de módulo de elasticidad de 10,4 Gpa en el ensayo de tracción para la trabécula ósea, mientras que en las muestras de hueso cortical se encuentra en 18,6 Gpa (21).
Los dos componentes del hueso tienen propiedades mecánicas extremadamente diferentes.
El mineral es duro y frágil, mientras que la proteína (húmeda) es mucho mas blanda pero más resistente (a la tracción, compresión, torsión, etc ).
Sin embargo la unión de ambos (composite) combina las propiedades óptimas de cada uno, aportando dureza y resistencia (21).
Un factor que va a condicionar las propiedades del hueso cortical o trabecular va a ser la proporción de placas minerales que contenga la matriz colagénica.
Se ha observado que después de un periodo de 10-15 días, la matriz colagénica inicia un proceso de mineralización primaria muy rápido (entre varios días y meses dependien...
| Tipo de Hueso | Módulo de Elasticidad (GPa) | Resistencia a la Tensión (MPa) | Resistencia a la Compresión (MPa) |
|---|---|---|---|
| Cortical | 15-20 | 80-150 | 90-280 |
| Trabecular | 0.75-20 (variable) | Variable | Variable |
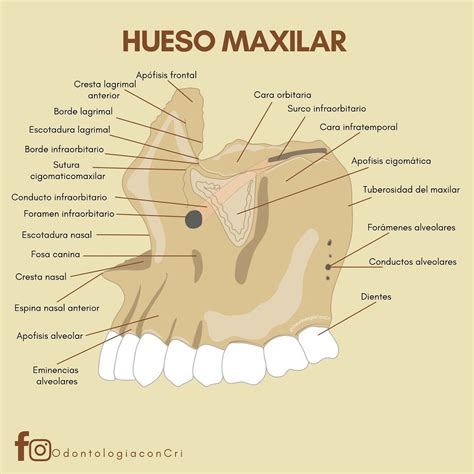
Anatomía del Hueso Maxilar
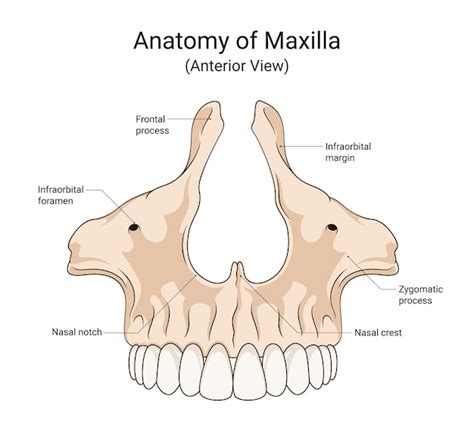
Vista Posterior del Maxilar